Биология - ВИЧ - Биология ВИЧ
09 февраля 2011Оглавление:
1. ВИЧ
2. История открытия
3. Биология ВИЧ
4. ВИЧ-инфекция
5. Эпидемиология
6. Анти-ВИЧ-кремы и гели
7. Юридические последствия заражения другого человека ВИЧ-инфекцией
Попадая в организм человека, ВИЧ заражает CD4+ лимфоциты, макрофаги и некоторые другие типы клеток. Проникнув же в указанные типы клеток, вирус начинает активно в них размножаться. Это в конечном счёте приводит к разрушению и гибели зараженных клеток. Присутствие ВИЧ со временем вызывает нарушение иммунной системы из-за избирательного уничтожения им иммунокомпетентных клеток и подавления их субпопуляции. Вышедшие из клетки вирусы внедряются в новые, и цикл повторяется. Постепенно число CD4+ лимфоцитов снижается настолько, что организм уже не может противостоять возбудителям оппортунистических инфекций, которые не опасны или мало опасны для здоровых людей с нормальной иммунной системой.
Классификация
Вирус иммунодефицита человека относят к семейству ретровирусов, роду лентивирусов. Название Lentivirus происходит от латинского слова lente — медленный. Такое название отражает одну из особенностей вирусов этой группы, а именно — медленную и неодинаковую скорость развития инфекционного процесса в макроорганизме. Для лентивирусов также характерен длительный инкубационный период.
Родственные вирусы
В роде Lentivirus выделяют следующие виды.
| Аббревиатура | Английское название | Русское название |
|---|---|---|
| EIAV | Equine infectious anemia virus | Вирус инфекционной анемии лошадей |
| OOP | Ovine Progressive Pneumonia | Вирус меди-висна овец |
| CAEV | Caprine-ovine arthritis-encephalitis virus | Вирус артрита-энцефалита коз и овец |
| BIV | Bovine immunodeficiency virus | Вирус иммунодефицита крупного рогатого скота |
| FIV | Feline immunodefitiency virus | Вирус иммунодефицита кошек |
| PLV | Puma lentivirus | Лентивирус пум |
| SIV | Simian immunedeficiency virus | Вирус иммунодефицита обезьян. Известно несколько штаммов этого вируса. Каждый штамм характерен для одного вида приматов: SIV-agm, SIV-cpz, SIV-mnd, SIV-mne, SIV-mac, SIV-sm, SIV-stm |
| HIV-1 | Human immunodeficiency virus-1 | Вирус иммунодефицита человека |
| HIV-2 | Human immunodeficiency virus-2 | Вирус иммунодефицита человека-2 |
Наиболее хорошо изученным является ВИЧ.
Разновидности ВИЧ
Для вируса иммунодефицита человека характерна высокая частота генетических изменений, возникающих в процессе самовоспроизведения. Частота возникновения ошибок у ВИЧ составляет 10 — 10 ошибок /, что на несколько порядков больше аналогичной величины у эукариот. Длина генома ВИЧ составляет примерно 10 нуклеотидов. Из этого следует, что практически каждый вирус хотя бы на один нуклеотид отличается от своего предшественника. В природе ВИЧ существует в виде множества квази-видов, являясь при этом одной таксономической единицей. В процессе исследования ВИЧ все-таки были обнаружены разновидности, которые значительно отличались друг от друга по нескольким признакам, в частности различной структурой генома. Разновидности ВИЧ обозначаются арабскими цифрами. На сегодняшний день известны ВИЧ-1, ВИЧ-2, ВИЧ-3, ВИЧ-4.
- ВИЧ-1 — первый представитель группы, открытый в 1983 году. Является наиболее распространенной формой.
- ВИЧ-2 — вид вируса иммунодефицита человека, идентифицированный в 1986 году . По сравнению с ВИЧ-1, ВИЧ-2 изучен в значительно меньшей степени. ВИЧ-2 отличается от ВИЧ-1 в структуре генома. Известно, что ВИЧ-2 менее патогенен и передается с меньшей вероятностью, чем ВИЧ-1. Отмечено, что люди, инфицированные ВИЧ-2, обладают слабым иммунитетом к ВИЧ-1.
- ВИЧ-3 — редкая разновидность, об открытии которой было сообщено в 1988. Обнаруженный вирус не реагировал с антителами других известных групп, а также обладал значительными отличиями в структуре генома. Более распространенное наименование для этой разновидности — ВИЧ-1 подтип O.
- ВИЧ-4 — редкая разновидность вируса, обнаруженная в 1986 году.
Глобальная эпидемия ВИЧ-инфекции главным образом обусловлена распространением ВИЧ-1. ВИЧ-2 распространен преимущественно в Западной Африке. ВИЧ-3 и ВИЧ-4 не играют заметной роли в распространении эпидемии.
В подавляющем большинстве случаев, если не оговорено иначе, под ВИЧ подразумевается ВИЧ-1.
Строение вириона
Вирионы ВИЧ имеют вид сферических частиц, диаметр которых составляет около 100—120 нанометров. Это приблизительно в 60 раз меньше диаметра эритроцита.
Капсид зрелого вириона имеет форму усеченного конуса. Иногда встречаются «многоядерные» вирионы, содержащие 2 или более нуклеоидов.
В состав зрелых вирионов входит несколько тысяч белковых молекул различных типов.
Названия и функции основных структурных белков ВИЧ-1.
| Сокращение | Описание | Функции |
|---|---|---|
| gp41 | трансмембранный гликопротеин массой 41 кДа | Располагается во внешнем слое липидной мембраны. Играет роль «якоря», удерживающего молекулы другого белка — gp120 |
| gp120 | гликопротеин массой 120 кДа | Наружный белок вириона. Нековалентно связан с трансмембранным белком gp41. С одной молекулой gp41 связаны 3 — 5 молекул gp120. Способен связываться с CD4 рецептором. Играет важную роль в процессе проникновения вируса в клетку. |
| p24 | белок массой 24 кДа | белок, образующий оболочку нуклеоида вируса |
| p17 | Матриксный белок массой 17 кДа | Около двух тысяч молекул этого белка образуют слой толщиной 5 — 7 нм, располагающийся между внешней оболочкой и нуклеоидом вируса. |
| p7 | Нуклеокапсидный белок массой 7 кДа | Белок, входящий в состав нуклеоида вируса. Образует комплекс с вирусной РНК. |
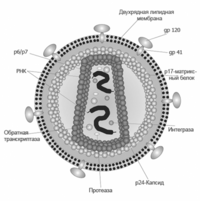
Внутри капсида ВИЧ находится белковонуклеиновый комплекс: две нити вирусной РНК, вирусные ферменты и белок p7. С капсидом также ассоциированы белки Nef и Vif. Внутри вириона обнаружен белок Vpr. Сам капсид образован ~2,000 копий вирусного белка p24. Стехиометрическое соотношение p24:gp120 в вирионе составляет 60-100:1, а p24:Pol примерно 10-20:1. Кроме того, с капсидом ВИЧ-1 связываются ~200 копий клеточного циклофилина А, который вирус заимствует у зараженной клетки .
Капсид ВИЧ окружен матриксной оболочкой, образованной ~2,000 копий матриксного белка p17. Матриксная оболочка в свою очередь окружена двуслойной липидной мембраной, являющейся наружной оболочкой вируса. Она образована молекулами, захваченными вирусом во время его отпочковывания из клетки, в которой он сформировался. В липидную мембрану встроены 72 гликопротеиновых комплекса, каждый из которых образован тремя молекулами трансмембранного гликопротеина, служащими «якорем» комплекса, и тремя молекулами поверхностного гликопротеина . С помощью gp120 вирус присоединяется к CD4 рецептору и корецептору, находящимся на поверхности мембраны клеток. gp41 и в особенности gp120 интенсивно изучаются как цели для разработки лекарств и вакцины против ВИЧ. В липидной мембране вируса также находятся мембранные белки клеток, в том числе человеческие лейкоцитарные антигены классов I, II и молекулы адгезии .
Геном ВИЧ-1 и кодируемые им белки
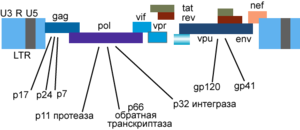
Генетический материал ВИЧ представлен двумя не связанными нитями положительно-смысловой) РНК . Геном ВИЧ-1 имеет длину 9,000 нуклеотидов. Концы генома представлены длинными концевыми повторами, которые управляют продукцией новых вирусов и могут активироваться и белками вируса, и белками инфицированной клетки.
9 генов ВИЧ-1 кодируют, по крайней мере, 15 белков .
- pol — кодирует ферменты: обратную транскриптазу, интегразу и протеазу.
- gag — кодирует полипротеин Gag/p55, расщепляемый вирусной протеазой до структурных белков p6, p7, p17, p24.
- env — кодирует белок gp160, расщепляемый клеточной эндопротеазой фурином на структурные белки gp41 и gp120.
Другие шесть генов — tat, rev, nef, vif, vpr, vpu — кодируют белки, отвечающие за способность ВИЧ-1 инфицировать клетки и производить новые копии вируса.
Репликация ВИЧ-1 in vitro возможна без генов nef, vif, vpr, vpu . Однако данные белки необходимы для полноценной инфекции in vivo.
Gag
Полипротеин-предшественник Gag/p55 синтезируется с полноразмерной геномной РНК в процессе стандартной кэп-зависимой трансляции, но синтез возможен и с помощью IRES, расположенного в 5'-нетранслируемой области мРНК. Локализация отдельных белков Gag, на которые расщепляется Gag/p55, следующая: p17...p24...p2...p7...p1...p6. Нерасщеплённый протеазой Gag/p55 содержит три основных домена: домен мембранной локализации, домен взаимодействия и "поздний" домен. Домен М, расположенный внутри области p17/МА, миристилируется и направляет Gag/p55 к плазматической мембране. Домен I, находящийся внутри области p7/NC, отвечает за межмолекулярные взаимодействия отдельных мономеров Gag/p55. Домен L, также локализованный в области p7/NC, опосредует отпочковывание дочерних вирионов от плазматической мембраны; в этом процессе участвует также р6 область полипротеина Gag/p55.
Vpu
Двумя важными функциями белка Vpu являются: 1) деградация клеточного рецептора CD4 в эндоплазматическом ретикулуме путём привлечения убиквитин-лигазных комплексов, и 2) стимуляция выделения дочерних вирионов из клетки, путём инактивации интерферон-индуцируемого трансмембранного белка CD317/BST-2, получившего также название «tetherin» за его способность подавлять выделение вновь образовавшихся дочерних вирионов посредством их удержания на поверхности клетки.
Vpr
Vif
Nef
Tat
Rev
Просмотров: 21058
|
|