Биология - Трансляция (биология) - Инициация
16 апреля 2011Оглавление:
1. Трансляция (биология)
2. Инициация
3. Элонгация
4. Терминация
Синтез белка в большинстве случаев начинается с AUG-кодона, кодирующего метионин. Этот кодон обычно называют стартовым или инициаторным. Инициация трансляции предусматривает узнавание рибосомой этого кодона и привлечение инициаторной аминоацил-тРНК. Для инициации трансляции необходимо также наличие определённых нуклеотидных последовательностей в районе стартового кодона. Немаловажная роль в защите 5'-конца мРНК принадлежит 5'-кэпу. Существование последовательности, отличающей стартовый AUG от внутренних совершенно необходимо, так как в противном случае инициация синтеза белка происходила бы хаотично на всех AUG-кодонах.
Процесс инициации обеспечивается специальными белками — факторами инициации.
Механизмы инициации трансляции у про- и эукариот существенно отличаются: прокариотические рибосомы потенциально способны находить стартовый AUG и инициировать синтез на любых участках мРНК, в то время как эукариотические рибосомы обычно присоединяются к мРНК в области кэпа и сканируют её в поисках стартового кодона.
У прокариот
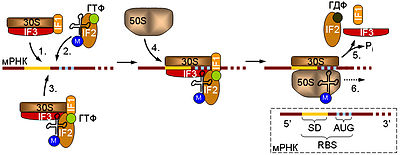
Начальная стадия предусматривает связывание малой рибосомной субъединицы с мРНК. Это может происходить двумя способами: либо сначала к мРНК присоединяется комплекс, содержащий рибосомную субчастицу, а затем к нему привлекается тРНК в комплексе с IF2 и ГТФ, либо 30S субъединица изначально связывается с тРНК, а уже потом садится на мРНК. К образовавшемуся комплексу приходит большая рибосомная субъединица, инициаторные факторы отсоединяются от 30S субчастицы, что сопровождается гидролизом ГТФ белком IF2, и собранная рибосома начинает элонгировать цепь. В правом нижнем углу дана схема инициаторного участка прокариотической мРНК. Отмечены 5' и 3' концы молекулы. RBS — сайт связывания рибосомы, SD — последовательность Шайн-Дальгарно, AUG — инициаторный кодон.
Малая рибосомная субъединица прокариот, если она не вовлечена в данный момент в трансляцию, существует в комплексе с инициаторными факторами IF1, IF3, и, в некоторых случаях, IF2. Рассмотрим основные функции этих белков:
- IF3, связанный с 30S-субъединицей, предотвращает ассоциацию с большой субъединицей рибосомы, тем самым сохраняя ее свободное состояние до связывания с матричной РНК. Этот белок также принимает участие в связывании мРНК и тРНК, а также IF2.
- IF2 взаимодействует с тРНК, а также обладает способностью расщеплять ГТФ.
- IF1 является, по-видимому, не обязательным фактором повышающим сродство малой субчастицы к IF2 и IF3.
Комплекс 30S субчастицы с инициаторными факторами способен узнавать специальные последовательности мРНК, так называемые участки связывания рибосомы. Эти участки содержат, во-первых, инициаторный AUG, и, во-вторых, специальную последовательность Шайна-Дальгарно с которой комплементарно связывается рибосомная 16S РНК. Последовательность Шайн-Дальгарно служит для того, чтобы отличать инициаторный AUG от внутренних кодонов, кодирующих метионин. После того, как 30S-субъединица связалась с мРНК к ней привлекается инициаторная аминоацил-тРНК и IF2, если они еще не были включены в комплекс. Затем присоединяется 50S-субчастица, происходит гидролиз ГТФ и диссоциация инициаторных факторов. Собранная рибосома начинает синтезировать полипептидную цепь.
У эукариот
У эукариот существуют два механизма нахождения рибосомой стартового AUG: кэп-зависимый и кэп-независимый.
- При сканирующем механизме рибосома садится на 5'-конец мРНК в области кэпа и двигается вдоль молекулы мРНК, «сканируя» один кодон за другим, пока не наткнётся на инициаторный AUG. Для привлечения рибосомы к 5'-концу мРНК требуется специальная структура, кэп — 7-метилгуанин, прикреплённый к 5'-концевому нуклеотиду мРНК.
- При механизме внутренней инициации, называемом у эукариот также IRES-зависимым механизмом, рибосома садится на внутренний участок мРНК, называемый IRES — участок мРНК, обладающий выраженной вторичной структурой, позволяющей ему направлять рибосомы на стартовый AUG. По IRES-зависимому механизму инициируется синтез лишь на небольшой части клеточных мРНК, а также на РНК некоторых вирусов .
Также у эукариот возможна реинициация трансляции, когда после окончания трансляции рибосома с белковыми факторами не диссоциирует от мРНК, а перескакивает с 3' на 5' конец мРНК и начинает инициацию ещё раз. Такое возможно благодаря замкнутой кольцевой форме мРНК в цитоплазме.
Кэп-зависимый механизм
В отличие от прокариот, инициация трансляции у которых обеспечивается лишь тремя белковыми факторами, трансляция подавляющего большинства мРНК эукариот, содержащих 5'-кэп и 3' поли-хвост, требует участия, по крайней мере, 13 общих эукариотических факторов инициации, представленных 31 полипептидом. Инициация трансляции включает события между диссоциацией рибосомы во время терминации в предыдущем цикле трансляции и сборкой рибосомы, готовой к элонгации, на старт-кодоне мРНК. Во время инициации аппарат трансляции решает следующие задачи:
- диссоциация и антиассоциация рибосомных субъединиц;
- выбор инициаторной метионил-тРНК;
- связывание 5'-кэпа, связывание поли, сканирование;
- выбор правильного старт-кодона;
- объединение рибосомных субъединиц на старт-кодоне
Диссоциация и антиассоциация субъединиц рибосом
Диссоциация рибосомных субъединиц в конце терминации — активный процесс, в котором участвуют eIF, а также факторы элонгации и терминации. Антиассоциация уже диссоциированных субъединиц обеспечивается eIF и служит для предотвращения преждевременного объединения рибосомных субъединиц. Главная роль в выполнении этой задачи принадлежит eIF3, мультисубъединичному фактору, состоящему из 13 различных субъединиц у млекопитающих, 11 субъединиц у растений и шести субъединиц у дрожжей Saccharomyces cerevisiae. eIF3 связывается с 40S субъединицей рибосомы посредством своей j-субъединицы, которая, в свою очередь, взаимодействует с «каркасной» b-субъединицей и предотвращает ассоциацию 40S с 60S рибосомной субъединицей. Эти активности eIF3 зависят от его взаимодействия с eIF1 и тройственным комплексом eIF2/GTP/Met-tRNAiMet. Связывание eIF1 с 40S является кооперативным с eIF3, так же как и связывание eIF1 с eIF1А. Таким образом, eIF1А, вероятно, также участвует в антиассоциации, по крайней мере, непрямым образом.
Cелекция инициаторной метионил-тРНК
Этот этап включает в себя следующие процессы:
- узнавание и метионилирование tRNAiMet специфичной метионил-тРНК-синтетазой;
- дискриминацию против Met-tRNAiMet эукариотическими факторами элонгации;
- дискриминацию против неметионилированной или неправильно аминоацилированной tRNAiMet eIF;
- дискриминацию против элонгаторных тРНК eIF.
В ходе процесса, метионил-тРНК-синтетаза взаимодействует как с акцепторным концом тРНК, так и с антикодоном.
Процесс у растений и дрожжей осуществляется с помощью посттранскрипционной модификации tRNAiMet, которая делает её отличной от элонгаторной метионин-специфичной тРНК с помощью присоединения 2'-О-фосфорибозила к рибозе нуклеотида А64. У позвоночных процесс осуществляется путём дискриминации между специфическими особенностями нуклеотидных последовательностей tRNAiMet и элонгаторной метиониновой тРНК.
Просмотров: 26501
|
|